
Mecanismos de interacción de los genes
También conocidos como interacción genética o interacción génica o simplemente acción génica. Son otros tipos de herencia, conocidos muchas veces como herencia no mendeliana, debido a que muchos de ellos no cumplen con las proporciones fenotípicas observadas por Mendel. Hay de dos tipos, interacciones entre alelos de un gen, o entre genes distintos, pero también aparecen otros tipos de herencia no mendeliana, como la pleiotropía, la penetrancia incompleta, los genes letales y otras formas de herencia distintas a las que describió Mendel, aquí incluimos la de los alelos múltiples que sí cumplen con las proporcione Mendelianas pero tienen otras particularidades.
Los mecanismos de interacción génica pueden clasificarse en los que ocurren entre alelos de un mismo gen (intra-alélicos) y los que se producen entre diferentes genes (inter-alélicos)
Intra-alélicos o entre alelos de un gen
Los intra alélicos o entre alelos de un mismo gen, son varios, uno ya mencionado en Mendel (la dominancia completa) y otros como la codominancia.
Son los siguientes:
- Dominancia completa
- Dominancia incompleta
- Codominancia
- Sobredominancia
Dominancia completa
Es el caso más descrito dónde un alelo domina o enmascara completamente a otro. Sin embargo es difícil explicar los fenómenos bioquímicos o moleculares que definen o generan este mecanismo; por lo tanto trataremos de explicarlo mediante ejemplos concretos.
El caso del alelo del albinismo (recesivo) ,tanto en humanos como en animales, es un ejemplo clásico de Dominancia Completa. Se cree que este alelo surgió como mutación del alelo que codifica o permite el fenotipo con pigmentación normal. Esto se da para la mayoría de las características, dónde el alelo normal o salvaje es dominante completo sobre el alelo mutante, en este caso recesivo.

El alelo normal (dominante) codifica alguna proteína quizás de función enzimática que interviene en la ruta biosintética para la formación de melanina.
Tanto los homocigotas dominantes como los heterocigotas tienen pigmentación normal. En el caso de los heterocigotas, producen melanina porque la existencia de un solo alelo normal o sea una sola copia funcional del gen, es suficiente para producir la enzima o proteína necesaria en esa ruta biosintética. En cambio los homocigotas recesivos, al no tener ningún alelo funcional, no producen producto y como consecuencia la síntesis de melanina es bloqueada con lo que los individuos carecen de la misma.A este tipo de alelo se lo denomina amorfo ya que es inactivo, no produciendo producto proteico alguno, ya sea producto de una mutación en la secuencia de DNA original o por la deleción del gen o parte del mismo.
Si denominamos «A» al alelo normal y «a» al productor del albinismo:
En algunos casos al revés de lo descripto antes, el alelo mutante domina sobre el salvaje o normal, como el caso del gen del enanismo y la acondroplasia en el hombre, donde los heterocigotas expresan el fenotipo alterado al igual que los homocigotas.
Existen muchas características cuyo mecanismo de acción entre alelos es la dominancia completa, sin embargo bioquímicamente son complicados de explicar. Por ejemplo la característica o enfermedad «Pie de Mula» en el Bovino, producida por un alelo recesivo, dando como resultado individuos que poseen un solo dedo como el equino. Esquemáticamente si denominamos «P» al alelo normal y «p» al afectado:
PP x pp Animal Normal apareado con uno con pié de Mula (pp)
F1: Pp 100% portadores del gen pero fenotípicamente normales
F1XF1: Pp x Pp
Proporción genotípica
1/4 PP (Homocigotas de fenotipo normal)
1/2 Pp (Heterocigotas o portadores de fenotipo normal)
1/4 pp Homocigotas recesivos (con pié de mula un sólo dedo como caballo)
Proporción fenotípica:
3/4 Individuos normales
1/4 Individuos con pié de mula
Como vemos siempre que exista Dominancia completa, las proporciones genotípicas y fenotípicas de la F2 , serán las descriptas por Mendel. Sin embargo sabiendo que un gen codifica una proteína, el pie de mula difícilmente se produzca por la falta o la alteración de una sola proteína. Creemos que deben ser varias las proteínas que intervengan en el desarrollo normal del pie Bovino, o quizás sea una sola proteína que intervenga en las primeras divisiones o segmentaciones del embrión, dando como resultado al estar alterada un fenotipo anormal.
Los alelos de un gen que por mutaciones en vez de convertirse en inactivos producen una proteína, pero alterada, como podría ser el pie de mula, se denominan hipomorfos.
Dominancia Incompleta
Carl Correns, uno de los redescubridores de Mendel cruzó experimentalmenteflores rojas con blancas, observando en la F1 la aparición de un nuevo fenotipo intermedio entre los parentales , es decir color rosa, lo que no coincidía con lo descripto por Mendel, ya que la F1 era uniforme pero no igual a uno de los progenitores.
A su vez al aparear a la F1 entre sí obtuvo una proporción fenotípica de 1/4 de flores rojas: ½ de flores rosas: ¼ de flores blancas. Si denominamos «R» al alelo del color rojo y «r» al blanco:
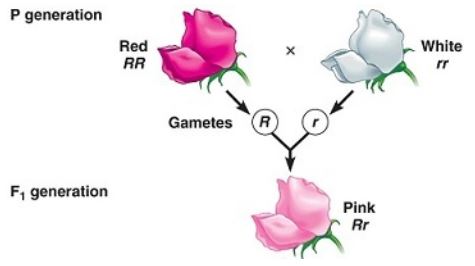
Al no existir dominancia entre un alelo y el otro, se prefiere usar como notación para los alelos, las letras mayúsculas pero con diferentes subíndices (R1 y R2). De esta forma no se indica si uno domina sobre el otro como usando mayúsculas y minúsculas. R1 R1 es el genotipo que expresa fenotipo rojo, R1 R2 rosa y el R2 R2 de fenotipo blanco.
En la actualidad si analizamos microscópicamente los cloroplastos de esas flores, observamos la existencia de cloroplastos rojos y blancos juntos, es decir que en forma macroscópica la flor se ve rosa pero microscópicamente tiene ambos colores, mecanismo llamado codominancia la que detallaremos a continuación.
En animales los casos descriptos son explicados hoy en día por la codominancia.
Codominancia
Es un mecanismo de acción entre alelos que se da cuando el heterocigota (F1) expresa fenotípicamente a ambos alelos de la característica en cuestión.
El ejemplo clásico es el del sistema de grupos sanguíneos ABO en los humanos. Los homocigotas para el alelo I A (IA IA) presentan o expresan fenotípicamente el grupo sanguíneo A, es decir antígenos en la superficie de los glóbulos rojos de tipo A. Los homocigotas para el alelo IB (IB IB), expresan fenotipo de grupo B, el heterocigota en cambio IA IB) tiene grupo AB es decir tiene ambos tipos de antígenos A y B.
Quiere decir que aquí tampoco un alelo domina sobre otro sino que se expresan ambos, tal como el ejemplo de las flores analizado anteriormente. Otros ejemplos en animales son el del pelaje rosillo en los equinos y bovinos, el que antes se explicaba como Dominancia Incompleta ya que en apariencia el rosillo era intermedio entre el colorado y el blanco, pero se sabe que es codominante ya que presenta pelos blancos y pelos colorados. Lo mismo ocurre en los gatos híbridos de siamés y burmese, que al ser heterocigotas presentan un fenotipo intermedio (tonquinés), pero en realidad se cree que expresan ambos fenotipos, las manchas del siamés oscuras y el pelaje pardo del burmese. Cabe aclarar que las manchas oscuras del siamés están influenciadas por la temperatura, ya que se presentan en zonas de piel fría, o sea que es un genotipo influenciado por el ambiente.


Este último es un ejemplo de codominancia y además de alelos múltiples ya que los grupos sanguíneos están codificados por un gen (I) que tiene tres variantes o alelos
Sobredominancia
Este tipo de interacción se da entre alelos de características cuantitativas (como el peso, altura, peso al destete, producción de leche, etc.), donde el heterocigota es superior numéricamente al promedio entre ambas líneas progenitoras homocigotas. Por ejemplo el peso al nacimiento de un individuo heterocigota será superior al peso al nacimiento promediado de los padres homocigotas. El caso será analizado como parte del vigor híbrido, el que se verá en cursos superiores.
Otros ejemplos en animales aquí

{kind=link}
¡Muchas gracias por esta información! Me sirvió mucho para mi tarea :3
De nada Meerii, gracias a vos por dejar estas palabras. Para esto es el blog!!!! me alegra mucho!! saludos
Hola Gabriela muy buena tu informacion la vd me ayuda mucho en mi tare me podrias ayudar con tu apellido por favor para un citado
Gracias Henry, claro mi nombre es Gabriela Marisa Iglesias. Gracias a vos por dejar un comentario
Hola Gabriela, una publicación que hizo encontré el siguiente ejercicio podría explicármelo, por favor.
1) En los gatos blancos, el color azulado del iris es un carácter dominante (A) con
respecto al color rojo (a). Determine el genotipo de todos los miembros de una familia
de gatos formada así: padre y madre blancos de ojos azules tienen un gatito de ojos
rojos, quien, cuando crece, se cruza con una gata de ojos azules y tienen, a su vez, un
cachorro con ojos rojos.
Hola Jula, siempre les explico que no es un sitio para obtener respuestas simples a preguntas hechas por otro docente ni para desarrollar ejercicios. Eso lo hago en clase. Es complejop para mí poder responder tantas preguntas de muchas personas, soy la única administradora del sitio. pero es claro que ambos padres portan un alelo recesivo para color de ojos. Saludos!!
Hola, tengo una consulta. El ejercicio plantea que dos razas puras de rana una azul y otra amarilla se cruzan, la progenie es toda azul. Pero en el cruzamiento de la progenie se obtiene individuos azules, amarillos y verdes en unas proporciones 7:7:2. No consigo averiguar qué tipo de herencia presenta este carácter. ¿Alguien podría ayudarme?
Gracias.
Hola Sergi, siempre les explico que no es un sitio para obtener respuestas simples a preguntas hechas por otro docente ni para desarrollar ejercicios. Eso lo hago en clase. Es complejop para mí poder responder tantas preguntas de muchas personas, soy la única administradora del sitio. Supongo a vuelo de pájaro, que quizás haya una epístasis allí…a pensar. Saludos
buenos días quisiera que me ayudara por favor con el siguiente ejercicio: en un cruzamiento se obtuvo la siguiente descendencia:
azul x escarlata 1/4 azul; 2/4 purpura; 1/4 escarlata
purpura x purpura 1/4 azul; 2/4 purpura; 1/4 escarlata
azul x azul 3/4 azul; 1/4 purpura
azul x purpura 3/8 azul; 4/8 purpura; 1/8 escarlata
purpura x escarlata 1/2 purpura; 1/2 escarlata
¿definir la relación de dominancia?
¿indicar el genotipo de los padres?
Muchas gracias
Hola Andrea, siempre les explico que no es un sitio para obtener respuestas simples a preguntas hechas por otro docente. No se que pretende el docente que que aprendas con eso pero, solo al voleo te puedo decir que tengas en cuenta que el cruzamiento que te plantean es similar al de la 2da ley de Mendel, pero con una diferencia importante. Especialmente en las proporciones fenotípicas, Lo ves?
Hola Gabriela, muy bueno el blog. ¡Muchas gracias!
Me gustaría preguntarte por qué se les denomina «intraalelicos» e «interalélicos», en vez de «intragénicos» e «intergénicos», a las excepciones de las leyes de Mendel. Me parecería más correcto esto último, o bien denominar «interalélicos» a codominancia, dominancia incompleta y alelismo múltiple por un lado, e «intergénicos» al resto.
Hola Rafa, estoy muy de acuerdo contigo, la verdad es que deberían ser llamados así como lo mencionas pero, como muchas veces, traté de usar la nomenclatura de los antiguos libros de genética. No me animo o al menos no me siento en condiciones de cambiarlos por mi antojo. Pero es así como deberían denominarse. Interalelicos e intergénicos. Saludos y gracias por comentar
¡Muchísimas gracias Gabriela por tu rápida respuesta! Me viene fenomenal de cara las oposiciones…glups,
Otra cosa: el cuadro donde se explica la epistasia recesiva tiene un pequeño error: aparece con fenotipo «chocolate» un genotipo EeBb, cuando ese debería ser negro (también aparece al revés).
Y con respecto a este fenómeno en concreto, el doble homocigótico recesivo tiene un fenotipo distinto al amarillo, lo cual cambia las frecuencias 9:3:4 a 9:3:3:1. ¿Estás de acuerdo? ¿Se consideran ambos ejemplos de epistasia recesiva?
Gracias otra vez
Rafa
Hola de nuevo Rafa y gracias por ser un poco mi editor en estas cosas. Ayuda mucho…a veces uno comente errores y nadie avisa, así que salud por ello. Ya lo he cambiado. Con respecto a lo de doble dominante y recesiva también a veces tengo discrepancias con la bibliografía y prefiero dar ejemplos sin ponerles mucho los nombres. Yo creo que sí debería tomarse como doble recesiva pero…en fin tu sabes. Si ves errores en otra página agradeceré tu ayuda. Saludos!!!!
Hola Gabriela! hace un tiempo pase por este blog para hacerte una consulta. vuelvo en este caso, para presentarte un problema que me dieron en un parcial y no pude encontrarle solución ya que desconocía el cambio en las proporciones.
me planteaban lo siguiente: cuando se cruza un hámster chino con manchas blancas con otro hámster chino sin manchas, aproximadamente un 1/2 de la progenie tiene manchas blancas y 1/2 no tiene manchas. cuando se cruzan dos hámsters con manchas blancas, 2/3 de las crías tiene manchas blancas y 1/3 carece de manchas.
a) ¿cuál es la base genética del manchado blanco en los hámster chinos?
b) ¿cómo prodrías producir hámsteres chinos con manchas blancas de raza pura?
Hola Rebeca siempre les explico que no es un sitio para obtener respuestas siemples a preguntas hechas por otro docente. No se que pretende el docente que que aprendas con eso pero, solo al voleo te puedo decir que leas en que son los genes letales y la dominancia y codominancia. Esta bien explicado en la pagina
Espero ayudar con eso
Saludossssssssssss
uff bueno lo lei y es tan simple como decir que el manchado blanco se debe a que el hamster con manchas blanca es heterocigoto, dominante solamente para manchas blancas y al cruzarse con uno sin manchas la matad sale sin manchas y la otra mitad con manchas. cuando se cruzan dos hamsters con machas blancas te das cuaenta una vez mas que el fenotipo de manchas blancas es heterocogoto de manera que si fueran homocigotos todos saldrian de manchas blancas. y para producir hamsters de manchas blancas de raza pura habria que eliminar cada descendiente sin manchas de cada descendencia asi se eliminarian los homocigotos resecivos de hamsters sin manchas 🙂
Gracias Jacqueline por tu respuesta y ayudarse entre Uds!!